
Plasmodesma - Plasmodesma

Plasmodesma (singular: plasmodesma ) são canais microscópicos que atravessam as paredes celulares das células vegetais e algumas células de algas , permitindo o transporte e a comunicação entre eles. Os plasmódios evoluíram independentemente em várias linhagens, e as espécies que possuem essas estruturas incluem membros das Charophyceae , Charales , Coleochaetales e Phaeophyceae (que são todas algas), bem como todos os embriófitos , mais conhecidos como plantas terrestres. Ao contrário das células animais , quase todas as células vegetais são rodeadas por um polissacarídeo parede celular . As células vegetais vizinhas são, portanto, separadas por um par de paredes celulares e a lamela intermediária intermediária , formando um domínio extracelular conhecido como apoplasto . Embora as paredes celulares sejam permeáveis a pequenas proteínas solúveis e outros solutos , os plasmodesmos permitem o transporte direto, regulado e simplástico de substâncias entre as células. Existem duas formas de plasmodesmos: plasmodos primários, que são formados durante a divisão celular, e plasmodesmas secundários, que podem se formar entre células maduras.
Estruturas semelhantes, chamadas de junções de hiato e nanotubos de membrana , interconectam as células animais e os estrômulos se formam entre os plastídios nas células vegetais.
Formação
Plasmodesmas primários são formados quando frações do retículo endoplasmático são aprisionadas através da lamela média à medida que uma nova parede celular é sintetizada entre duas células vegetais recém-divididas. Essas eventualmente se tornam as conexões citoplasmáticas entre as células. No local da formação, a parede não é mais espessada, e depressões ou áreas delgadas conhecidas como fossos são formadas nas paredes. Os poços normalmente formam pares entre as células adjacentes. Os plasmódios também podem ser inseridos nas paredes celulares existentes entre as células que não se dividem (plasmodesmos secundários).
Plasmodesma primário
A formação dos plasmódios primários ocorre durante a parte do processo de divisão celular onde o retículo endoplasmático e a nova placa se fundem, esse processo resulta na formação de um poro citoplasmático (ou manga citoplasmática). O desmotúbulo, também conhecido como ER comprimido, forma-se ao lado do ER cortical. Tanto o ER comprimido quanto o ER cortical são compactados firmemente juntos, não deixando espaço para nenhum espaço luminal. É proposto que o ER comprimido atua como uma rota de transporte de membrana nos plasmodos. Quando os filamentos do RE cortical são emaranhados na formação de uma nova placa de células, a formação de plasmodesma ocorre em plantas terrestres. É hipotetizado que o ER comprimido se forma devido a uma combinação de pressão de uma parede celular em crescimento e interação das proteínas ER e PM. Plasmodesmas primários estão freqüentemente presentes em áreas onde as paredes celulares parecem ser mais finas. Isso se deve ao fato de que, à medida que a parede celular se expande, a abundância dos plasmódios primários diminui. A fim de expandir ainda mais a densidade plasmódica durante o crescimento da parede celular, são produzidos plasmodesmos secundários. O processo de formação de plasmodesmas secundários ainda não foi totalmente compreendido, no entanto, várias enzimas degradantes e proteínas ER são consideradas como estimulantes do processo.
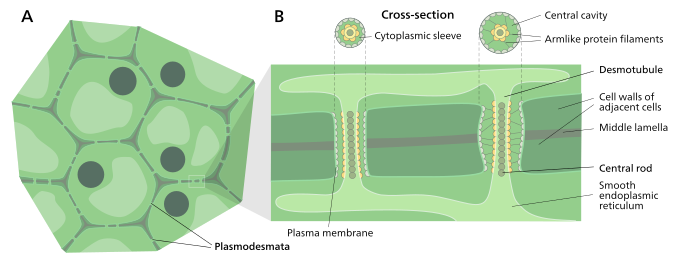
Estrutura
Membrana plasmática plasmática
Uma célula de planta típica pode ter entre 10 3 e 10 5 plasmodesmas conectando-a com células adjacentes igualando entre 1 e 10 por µm 2 . Os plasmódios têm aproximadamente 50-60 nm de diâmetro no ponto médio e são constituídos por três camadas principais, a membrana plasmática , a manga citoplasmática e o desmotúbulo . Eles podem atravessar paredes celulares de até 90 nm de espessura.
A porção da membrana plasmática do plasmodesma é uma extensão contínua da membrana celular ou plasmalema e tem uma estrutura de bicamada fosfolipídica semelhante .
A manga citoplasmática é um espaço preenchido por fluido delimitado pelo plasmalema e é uma extensão contínua do citosol . O tráfico de moléculas e íons através dos plasmodesmos ocorre neste espaço. Moléculas menores (por exemplo, açúcares e aminoácidos ) e íons podem facilmente passar pelos plasmódios por difusão sem a necessidade de energia química adicional. Moléculas maiores, incluindo proteínas (por exemplo, proteína verde fluorescente ) e RNA, também podem passar difusivamente pela manga citoplasmática. O transporte plasmático de algumas moléculas maiores é facilitado por mecanismos atualmente desconhecidos. Um mecanismo de regulação da permeabilidade dos plasmodesmos é o acúmulo do polissacarídeo caloso em torno da região do pescoço para formar um colar, reduzindo assim o diâmetro do poro disponível para o transporte das substâncias. Por meio da dilatação, passagem ativa ou remodelação estrutural, a permeabilidade dos plasmodesmatos é aumentada. Esse aumento na permeabilidade dos poros dos plasmodos permite que moléculas maiores, ou macromoléculas , como moléculas de sinalização, fatores de transcrição e complexos RNA-proteína sejam transportados para vários compartimentos celulares.
Estrutura de um plasmodesma e sua localização dentro das células vegetais

Os plasmódios permitem que as moléculas viajem entre as células vegetais através da via simplástica


Desmotúbulo
O desmotubule é um tubo de apresso (achatada) retículo endoplasmático que corre entre duas células adjacentes. Sabe-se que algumas moléculas são transportadas por esse canal, mas não se acredita que seja a principal via de transporte plasmodesmatal.
Ao redor do desmotúbulo e da membrana plasmática foram vistas áreas de um material denso de elétrons, muitas vezes unidas por estruturas semelhantes a raios que parecem dividir o plasmodesma em canais menores. Essas estruturas podem ser compostas de miosina e actina , que fazem parte do citoesqueleto da célula . Se for esse o caso, essas proteínas poderiam ser usadas no transporte seletivo de grandes moléculas entre as duas células.
Transporte

Foi demonstrado que os plasmódios transportam proteínas (incluindo fatores de transcrição ), RNA de interferência curto , RNA mensageiro , viróides e genomas virais de célula para célula. Um exemplo de proteínas de movimento viral é o vírus do mosaico do tabaco MP-30. Pensa-se que o MP-30 se liga ao próprio genoma do vírus e o transporta das células infectadas para as células não infectadas através dos plasmodos. A proteína do Locus T de floração se move das folhas para o meristema apical do caule através dos plasmodesmos para iniciar a floração .
Os plasmódios também são usados pelas células no floema , e o transporte simplástico é usado para regular as células do tubo da peneira pelas células companheiras .
O tamanho das moléculas que podem passar pelos plasmódios é determinado pelo limite de exclusão de tamanho. Este limite é altamente variável e está sujeito a modificações ativas. Por exemplo, o MP-30 é capaz de aumentar o limite de exclusão de tamanho de 700 Daltons para 9400 Daltons, ajudando assim seu movimento através de uma planta. Além disso, o aumento das concentrações de cálcio no citoplasma, seja por injeção ou por indução a frio, mostrou restringir a abertura dos plasmódios circundantes e limitar o transporte.
Existem vários modelos para possível transporte ativo através dos plasmódios. Foi sugerido que esse transporte é mediado por interações com proteínas localizadas no desmotúbulo e / ou por proteínas parcialmente desdobradas de chaperones , permitindo que elas se encaixem na passagem estreita. Um mecanismo semelhante pode estar envolvido no transporte de ácidos nucléicos virais através dos plasmodos.
Vários modelos matemáticos foram sugeridos para estimar o transporte entre os plasmódios. Esses modelos trataram principalmente o transporte como um problema de difusão com alguns obstáculos adicionais.
Componentes do citoesqueleto de plasmódios
Os plasmódios ligam quase todas as células de uma planta, o que pode causar efeitos negativos, como a disseminação de vírus. Para entender isso, devemos primeiro olhar para os componentes do citoesqueleto, como microfilamentos de actina, microtúbulos e proteínas de miosina, e como eles estão relacionados ao transporte de célula para célula. Os microfilamentos de actina estão ligados ao transporte de proteínas do movimento viral para os plasmódios, o que permite o transporte de célula para célula através dos plasmodos. A marcação fluorescente para co-expressão em folhas de tabaco mostrou que os filamentos de actina são responsáveis pelo transporte de proteínas de movimento viral para os plasmódios. Quando a polimerização da actina foi bloqueada, ela causou uma diminuição no direcionamento dos plasmódios das proteínas de movimento no tabaco e permitiu que componentes de 10 kDa (em vez de 126 kDa) se movessem entre as células do mesofilo do tabaco . Isso também afetou o movimento de célula para célula de moléculas dentro da planta do tabaco.
Vírus
Os vírus quebram os filamentos de actina dentro do canal do plasmodesma para se moverem dentro da planta. Por exemplo, quando o vírus do mosaico do pepino (CMV) chega às plantas, ele é capaz de viajar através de quase todas as células por meio da utilização de proteínas de movimento viral para se transportar através dos plasmodos. Quando as folhas de tabaco são tratadas com uma droga que estabiliza os filamentos de actina, a faloidina, as proteínas de movimento do vírus do mosaico do pepino são incapazes de aumentar o limite de exclusão de tamanho dos plasmódios (SEL).
Miosina
Grandes quantidades de proteínas de miosina são encontradas nos locais dos plasmodos. Essas proteínas estão envolvidas no direcionamento de cargas virais para os plasmódios. Quando as formas mutantes de miosina foram testadas em plantas de tabaco, o direcionamento da proteína viral aos plasmódios foi afetado negativamente. A ligação permanente da miosina à actina, que foi induzida por uma droga, causou uma diminuição no movimento de célula para célula. Os vírus também são capazes de se ligar seletivamente às proteínas da miosina.
Microtúbulos
Os microtúbulos têm um papel importante no transporte de RNA viral de célula para célula. Os vírus usam muitos métodos diferentes para se transportar de célula para célula e um desses métodos associa o domínio N-terminal de seu RNA para se localizar em plasmódios por meio de microtúbulos. Em plantas de tabaco injetadas com vírus do mosaico do tabaco que foram mantidas em altas temperaturas, houve uma forte correlação entre as proteínas de movimento TMV marcadas com GFP e os microtúbulos. Isso levou a um aumento na disseminação do RNA viral através do tabaco.
Plasmódico e caloso
A regulação e a estrutura dos plasmódios são reguladas por um polímero beta 1,3-glucano conhecido como caloso. Calose é encontrada em placas de células durante o processo de citocinese, mas à medida que esse processo atinge a conclusão, os níveis de calosidade diminuem. As únicas partes ricas e calosas da célula incluem as seções da parede celular onde os plasmódios estão presentes. Para regular o que é transportado pelos plasmodesmos, deve haver calosidade. Callose fornece o mecanismo pelo qual a permeabilidade do plasmodesma é regulada. A fim de controlar o que é transportado entre diferentes tecidos, os plasmodesmos sofrem várias alterações conformacionais especializadas.
A atividade dos plasmodesmas está ligada a processos fisiológicos e de desenvolvimento dentro das plantas. Existe uma via de sinalização hormonal que retransmite os sinais celulares primários através dos plasmodesmos. Existem também padrões de pistas ambientais, fisiológicas e de desenvolvimento que mostram relação com a função dos plasmódios. Um mecanismo importante dos plasmodesmas é a capacidade de bloquear seus canais. Os níveis de calosidade provaram ser um método para alterar o tamanho da abertura dos plasmodos. Depósitos calosos são encontrados no pescoço dos plasmodesmos em novas paredes celulares que foram formadas. O nível de depósitos nos plasmódios pode flutuar, o que mostra que há sinais que desencadeiam um acúmulo de calosidade nos plasmódios e fazem com que os plasmódios se tornem fechados ou mais abertos. As atividades enzimáticas da beta 1,3-glucano sintase e das hidrolases estão envolvidas nas alterações no nível de celulose dos plasmodos. Alguns sinais extracelulares alteram a transcrição das atividades desta sintase e da hidrolase. Arabidopsis thaliana tem genes de sintase calosa que codificam uma subunidade catalítica de B-1,3-glucano. Os mutantes de ganho de função neste pool de genes mostram um aumento da deposição de calosidade nos plasmodos e uma diminuição no tráfego macromolecular, bem como um sistema radicular defeituoso durante o desenvolvimento.