
Esfingomielina - Sphingomyelin

A esfingomielina ( SPH , ˌsfɪŋɡoˈmaɪəlɪn) é um tipo de esfingolipídeo encontrado nas membranas das células animais , especialmente na bainha de mielina membranosa que envolve alguns axônios das células nervosas . Geralmente consiste em fosfocolina e ceramida , ou um grupo de cabeça de fosfoetanolamina ; portanto, as esfingomielinas também podem ser classificadas como esfingofosfolipídios. Em humanos, o SPH representa ~ 85% de todos os esfingolipídios e, normalmente, compõe 10–20 mol% dos lipídios da membrana plasmática .
A esfingomielina foi isolada pela primeira vez pelo químico alemão Johann LW Thudicum na década de 1880. A estrutura da esfingomielina foi relatada pela primeira vez em 1927 como N-acil-esfingosina-1-fosforilcolina. O conteúdo de esfingomielina em mamíferos varia de 2 a 15% na maioria dos tecidos, com concentrações mais altas encontradas em tecidos nervosos, glóbulos vermelhos e lentes oculares. A esfingomielina tem papéis estruturais e funcionais significativos na célula. É um componente da membrana plasmática e participa de várias vias de sinalização. O metabolismo da esfingomielina cria muitos produtos que desempenham papéis significativos na célula.
Características físicas


Composição
A esfingomielina consiste em um grupo principal de fosfocolina , uma esfingosina e um ácido graxo . É um dos poucos fosfolipídios de membrana não sintetizados a partir do glicerol. A esfingosina e o ácido graxo podem ser categorizados coletivamente como ceramida. Esta composição permite que a esfingomielina desempenhe papéis significativos nas vias de sinalização: a degradação e a síntese da esfingomielina produzem segundos mensageiros importantes para a transdução do sinal.
A esfingomielina obtida de fontes naturais, como ovos ou cérebro bovino, contém ácidos graxos de vários comprimentos de cadeia. A esfingomielina com comprimento de cadeia definido, como palmitoilsfingomielina com uma cadeia de 16 acil saturada, está disponível comercialmente.
Propriedades
Idealmente, as moléculas de esfingomielina têm a forma de um cilindro, no entanto, muitas moléculas de esfingomielina têm uma incompatibilidade de cadeia significativa (os comprimentos das duas cadeias hidrofóbicas são significativamente diferentes). As cadeias hidrofóbicas da esfingomielina tendem a ser muito mais saturadas do que outros fosfolipídios. A principal temperatura de fase de transição das esfingomielinas também é maior em comparação à temperatura de transição de fase de fosfolipídios semelhantes, próxima a 37 ° C. Isso pode introduzir heterogeneidade lateral na membrana, gerando domínios na bicamada da membrana.
A esfingomielina sofre interações significativas com o colesterol. O colesterol tem a capacidade de eliminar a transição da fase líquida para a fase sólida nos fosfolipídios. Devido à temperatura de transição da esfingomielina estar dentro das faixas de temperatura fisiológica, o colesterol pode desempenhar um papel significativo na fase da esfingomielina. A esfingomielina também é mais propensa a ligações de hidrogênio intermoleculares do que outros fosfolipídios.
Localização
A esfingomielina é sintetizada no retículo endoplasmático (RE), onde pode ser encontrada em pequenas quantidades, e no trans Golgi. É enriquecido na membrana plasmática com uma concentração maior no folheto externo do que no interno. O complexo de Golgi representa um intermediário entre o RE e a membrana plasmática, com concentrações ligeiramente mais altas para o lado trans.
Metabolismo
Síntese
A síntese de esfingomielina envolve a transferência enzimática de uma fosfocolina da fosfatidilcolina para uma ceramida. A primeira etapa comprometida da síntese de esfingomielina envolve a condensação de L-serina e palmitoil-CoA . Esta reação é catalisada pela serina palmitoiltransferase . O produto dessa reação é reduzido, originando diidroesfingosina. A dihidroesfingosina sofre N-acilação seguida de dessaturação para produzir uma ceramida. Cada uma dessas reações ocorre na superfície citosólica do retículo endoplasmático . A ceramida é transportada para o aparelho de Golgi, onde pode ser convertida em esfingomielina. A esfingomielina sintase é responsável pela produção de esfingomielina a partir da ceramida. O diacilglicerol é produzido como um subproduto quando a fosfocolina é transferida.

Degradação
A quebra da esfingomielina é responsável por iniciar muitas vias universais de sinalização. É hidrolisado por esfingomielinases (fosfolipases tipo C específicas da esfingomielina). O grupo da cabeça da fosfocolina é liberado no meio aquoso enquanto a ceramida se difunde através da membrana.
Função
Membranas
A bainha de mielina membranosa que envolve e isola eletricamente muitos axônios das células nervosas é particularmente rica em esfingomielina, sugerindo seu papel como isolante das fibras nervosas. A membrana plasmática de outras células também é abundante em esfingomielina, embora seja amplamente encontrada no folheto exoplasmático da membrana celular. Há, entretanto, algumas evidências de que também pode haver um pool de esfingomielina no folheto interno da membrana. Além disso, a esfingomielinase-2 neutra - uma enzima que quebra a esfingomielina em ceramida - foi encontrada para se localizar exclusivamente no folheto interno, sugerindo ainda que pode haver esfingomielina presente lá.
Transdução de sinal
A função da esfingomielina permaneceu obscura até que foi descoberto que ela desempenha um papel na transdução do sinal . Foi descoberto que a esfingomielina desempenha um papel significativo nas vias de sinalização celular. A síntese de esfingomielina na membrana plasmática pela esfingomielina sintase 2 produz diacilglicerol, que é um segundo mensageiro lipossolúvel que pode passar ao longo de uma cascata de sinal. Além disso, a degradação da esfingomielina pode produzir ceramida que está envolvida na via de sinalização apoptótica.
Apoptose
Verificou-se que a esfingomielina desempenha um papel na apoptose celular por hidrólise em ceramida. Estudos no final da década de 1990 descobriram que a ceramida era produzida em uma variedade de condições que levavam à apoptose. Foi então levantada a hipótese de que a hidrólise da esfingomielina e a sinalização da ceramida eram essenciais na decisão sobre a morte de uma célula. No início dos anos 2000, surgiram novos estudos que definiram um novo papel para a hidrólise da esfingomielina na apoptose, determinando não apenas quando uma célula morre, mas como. Após mais experimentação, foi demonstrado que se a hidrólise da esfingomielina acontecer em um ponto suficientemente precoce na via, a produção de ceramida pode influenciar a taxa e a forma de morte celular ou trabalhar para liberar blocos em eventos a jusante.
Jangadas de lipídios
A esfingomielina, assim como outros esfingolipídeos, estão associados a microdomínios lipídicos na membrana plasmática conhecidos como jangadas lipídicas . As jangadas lipídicas são caracterizadas por as moléculas de lipídios estarem na fase ordenada de lipídios, oferecendo mais estrutura e rigidez em comparação com o resto da membrana plasmática. Nas jangadas, as cadeias de acila têm baixo movimento de cadeia, mas as moléculas têm alta mobilidade lateral. Essa ordem se deve em parte à alta temperatura de transição dos esfingolipídeos, bem como às interações desses lipídeos com o colesterol. O colesterol é uma molécula não polar relativamente pequena que pode preencher o espaço entre os esfingolipídeos, que é o resultado das grandes cadeias de acila. Acredita-se que as jangadas de lipídios estejam envolvidas em muitos processos celulares, como triagem e tráfego de membrana, transdução de sinal e polarização celular. O excesso de esfingomielina em jangadas lipídicas pode levar à resistência à insulina .
Devido aos tipos específicos de lipídios nesses microdomínios, os lipid rafts podem acumular certos tipos de proteínas a eles associados, aumentando, assim, as funções especiais que possuem. Especula-se que as jangadas lipídicas estão envolvidas na cascata da apoptose celular.
Anormalidades e doenças associadas
A esfingomielina pode se acumular em uma doença hereditária rara chamada doença de Niemann-Pick , tipos A e B. É uma doença herdada geneticamente causada por uma deficiência da enzima lisossomal esfingomielinase ácida , que causa o acúmulo de esfingomielina no baço , fígado , pulmões , medula óssea e cérebro , causando danos neurológicos irreversíveis. Dos dois tipos que envolvem esfingomielinase , o tipo A ocorre em bebês. É caracterizada por icterícia , fígado aumentado e danos cerebrais profundos . As crianças com este tipo raramente vivem além dos 18 meses. O tipo B envolve fígado e baço aumentados, o que geralmente ocorre na pré-adolescência. O cérebro não é afetado. A maioria dos pacientes apresenta níveis normais da enzima <1% em comparação aos níveis normais. Uma proteína hemolítica, lisenina, pode ser uma sonda valiosa para a detecção de esfingomielina em células de pacientes Niemann-Pick A.
Como resultado da doença autoimune esclerose múltipla (EM), a bainha de mielina das células neuronais no cérebro e na medula espinhal é degradada, resultando na perda da capacidade de transdução de sinal. Pacientes com esclerose múltipla exibem regulação positiva de certas citocinas no líquido cefalorraquidiano, particularmente fator de necrose tumoral alfa . Isso ativa a esfingomielinase, uma enzima que catalisa a hidrólise da esfingomielina em ceramida; A atividade da esfingomielinase foi observada em conjunto com a apoptose celular.
Um excesso de esfingomielina na membrana dos glóbulos vermelhos (como na abetalipoproteinemia ) causa acúmulo excessivo de lipídios no folheto externo da membrana plasmática dos glóbulos vermelhos . Isso resulta em glóbulos vermelhos de formato anormal chamados acantócitos .
Imagens adicionais

Modelo bola e bastão de esfingomielina
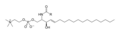
Fórmula esquelética de esfingomielina

Referências
links externos
- Esfingomielinas nos cabeçalhos de assuntos médicos da Biblioteca Nacional de Medicina dos EUA (MeSH)