
Aminoácidos não proteinogênicos - Non-proteinogenic amino acids

Em bioquímica , os aminoácidos não codificados ou não proteinogênicos são distintos dos 22 aminoácidos proteinogênicos (21 em eucariotos) que são codificados naturalmente no genoma dos organismos para a montagem de proteínas. No entanto, mais de 140 aminoácidos não proteinogênicos ocorrem naturalmente nas proteínas e milhares mais podem ocorrer na natureza ou ser sintetizados em laboratório. Muitos aminoácidos não proteinogênicos são importantes:
- intermediários na biossíntese,
- na formação pós-tradução de proteínas,
- em um papel fisiológico (por exemplo, componentes das paredes celulares bacterianas , neurotransmissores e toxinas ),
- compostos farmacológicos naturais ou artificiais,
- presente em meteoritos ou usado em experimentos pré-bióticos (por exemplo, experimento de Miller-Urey ).
Definição por negação

Tecnicamente, qualquer composto orgânico com uma amina (-NH 2 ) e um grupo funcional de ácido carboxílico (-COOH) é um aminoácido. Os aminoácidos proteinogênicos são um pequeno subconjunto deste grupo que possui átomo de carbono central (α- ou 2-) carregando um grupo amino, um grupo carboxila, uma cadeia lateral e uma conformação levo α-hidrogênio , com exceção da glicina , que é aquiral e prolina , cujo grupo amina é uma amina secundária e, consequentemente, é frequentemente referido como um iminoácido por razões tradicionais, embora não seja um imino.
O código genético codifica 20 aminoácidos padrão para incorporação em proteínas durante a tradução . No entanto, existem dois aminoácidos proteinogênicos extras: selenocisteína e pirrolisina . Estes aminoácidos não padrão não têm um códon dedicado, mas são adicionados no lugar de um códon de parada quando uma sequência específica está presente, códon UGA e elemento SECIS para selenocisteína, sequência UAG PYLIS a jusante para pirrolisina. Todos os outros aminoácidos são denominados "não proteinogênicos".

Selenocisteína. Este aminoácido contém um grupo selenol em seu carbono β

Pirrolisina. Este aminoácido é formado pela união ao grupo ε-amino da lisina de um anel de pirrolina carboxilado

Existem vários grupos de aminoácidos:
- 20 aminoácidos padrão
- 22 aminoácidos proteinogênicos
- mais de 80 aminoácidos criados abioticamente em altas concentrações
- cerca de 900 são produzidos por vias naturais
- mais de 118 aminoácidos projetados foram colocados em proteínas
Esses grupos se sobrepõem, mas não são idênticos. Todos os 22 aminoácidos proteinogênicos são biossintetizados por organismos e alguns, mas não todos, também são abióticos (encontrados em experimentos pré-bióticos e meteoritos). Alguns aminoácidos naturais, como a norleucina , são incorporados erroneamente à tradução em proteínas devido à infidelidade do processo de síntese de proteínas. Muitos aminoácidos, como a ornitina , são intermediários metabólicos produzidos biossinteticamente, mas não incorporados de forma translacional às proteínas. A modificação pós-tradução de resíduos de aminoácidos em proteínas leva à formação de muitos aminoácidos proteicos, mas não proteinogênicos. Outros aminoácidos são encontrados apenas em misturas abióticas (por exemplo, α-metilnorvalina). Mais de 30 aminoácidos não naturais foram inseridos de forma translacional em proteínas em sistemas projetados, mas não são biossintéticos.
Nomenclatura
Além do sistema de numeração IUPAC para diferenciar os vários carbonos em uma molécula orgânica, atribuindo sequencialmente um número a cada carbono, incluindo aqueles que formam um grupo carboxílico, os carbonos ao longo da cadeia lateral de aminoácidos também podem ser marcados com letras gregas , onde o carbono α é o carbono quiral central possuindo um grupo carboxila, uma cadeia lateral e, em α-aminoácidos, um grupo amino - o carbono em grupos carboxílicos não é contado. (Consequentemente, os nomes IUPAC de muitos α-aminoácidos não proteinogênicos começam com 2-amino e terminam em ácido in- ico .)
Natural, mas não L-α-aminoácidos
A maioria dos aminoácidos naturais são α-aminoácidos na conformação L, mas existem algumas exceções.
Não alfa

Alguns aminoácidos não-α existem em organismos. Nessas estruturas, o grupo amina se deslocou ainda mais da extremidade do ácido carboxílico da molécula de aminoácido. Assim, um aminoácido β tem o grupo amina ligado ao segundo carbono afastado, e um aminoácido γ o possui no terceiro. Os exemplos incluem β-alanina , GABA e ácido δ- aminolevulínico .

β-alanina: um aminoácido produzido pela aspartato 1-descarboxilase e um precursor da coenzima A e dos peptídeos carnosina e anserina .

Ácido γ-aminobutírico (GABA): um neurotransmissor em animais.

Ácido δ-aminolevulínico: um intermediário na biossíntese de tetrapirrol ( heme , clorofila , cobalamina etc. ).

Ácido 4-aminobenzoico (PABA): um intermediário na biossíntese de folato



A razão pela qual os α-aminoácidos são usados em proteínas tem sido associada à sua frequência em meteoritos e experimentos pré-bióticos. Uma especulação inicial sobre as propriedades deletérias dos β-aminoácidos em termos de estrutura secundária revelou-se incorreta.
D-aminoácidos
Alguns aminoácidos contêm a quiralidade absoluta oposta, substâncias químicas que não estão disponíveis na maquinaria de tradução / transcrição ribossomal normal. A maioria das paredes das células bacterianas é formada por peptidoglicano , um polímero composto de amino açúcares reticulados com oligopeptídeos curtos interligados. O oligopeptídeo é sintetizado não ribossomalmente e contém várias peculiaridades, incluindo D-aminoácidos , geralmente D-alanina e D-glutamato. Uma outra peculiaridade é que o primeiro é racemizado por uma enzima de ligação a PLP (codificada por alr ou o homólogo dadX ), enquanto o último é racemizado por uma enzima independente de cofator ( murI ). Algumas variantes estão presentes, em Thermotoga spp. A D-lisina está presente e em certas bactérias resistentes à vancomicina a D-serina está presente ( gene vanT ).
Em animais, alguns D-aminoácidos são neurotransmissores.
Sem um hidrogênio no carbono α
Todos os aminoácidos proteinogênicos têm pelo menos um hidrogênio no carbono α. A glicina tem dois hidrogênios e todos os outros têm um hidrogênio e uma cadeia lateral. A substituição do hidrogênio restante por um substituinte maior, como um grupo metil, distorce a estrutura da proteína.
Em alguns fungos, o ácido α-aminoisobutírico é produzido como um precursor de peptídeos, alguns dos quais exibem propriedades antibióticas. Este composto é semelhante à alanina, mas possui um grupo metil adicional no carbono α em vez de um hidrogênio. Portanto, é aquiral. Outro composto semelhante à alanina sem um α-hidrogênio é a desidroalanina , que possui uma cadeia lateral de metileno. É um dos vários desidroaminoácidos que ocorrem naturalmente .

alanina

ácido aminoisobutírico

desidroalanina



Estereocentros gêmeos de aminoácidos
Um subconjunto de L-α-aminoácidos é ambíguo quanto a qual das duas extremidades é o carbono α. Em proteínas, um resíduo de cisteína pode formar uma ligação dissulfeto com outro resíduo de cisteína, reticulando assim a proteína. Duas cisteínas reticuladas formam uma molécula de cistina . A cisteína e a metionina são geralmente produzidos por sulfurylation directa, mas em algumas espécies que podem ser produzidos por transulfuração , onde o activado homoserina ou serina é fundido com uma cisteína ou homocisteína formando cistationina . Um composto semelhante é a lantionina , que pode ser vista como duas moléculas de alanina unidas por uma ligação tioéter e é encontrada em vários organismos. Da mesma forma, o ácido djenkólico , uma toxina vegetal dos grãos jengkol , é composto de duas cisteínas conectadas por um grupo metileno. O ácido diaminopimélico é usado como uma ponte no peptidoglicano e é usado como um precursor da lisina (por meio de sua descarboxilação).

cistina
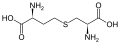
cistationina

lantionina
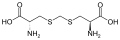
Ácido Djenkolic
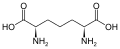
Ácido diaminopimélico

Aminoácidos prebióticos e bioquímicas alternativas
Em meteoritos e em experimentos pré-bióticos (por exemplo, experimento de Miller-Urey ), muitos mais aminoácidos do que os vinte aminoácidos padrão são encontrados, vários dos quais em concentrações mais altas do que as padrão: foi conjecturado que se a vida baseada em aminoácidos surgisse paralelamente, em outras partes do universo, não mais do que 75% dos aminoácidos seriam em comum. A anomalia mais notável é a falta de ácido aminobutírico.
| Proporção de aminoácidos em relação à glicina (%) | ||
|---|---|---|
| Molécula | Descarga elétrica | Meteorito Murchinson |
| glicina | 100 | 100 |
| alanina | 180 | 36 |
| ácido α-amino-n-butírico | 61 | 19 |
| norvalina | 14 | 14 |
| valina | 4,4 | |
| norleucina | 1,4 | |
| leucina | 2,6 | |
| isoleucina | 1,1 | |
| aloisoleucina | 1,2 | |
| t-leucina | <0,005 | |
| ácido α-amino-n-heptanoico | 0,3 | |
| prolina | 0,3 | 22 |
| ácido pipecólico | 0,01 | 11 |
| ácido α, β-diaminopropiônico | 1,5 | |
| ácido α, γ-diaminobutírico | 7,6 | |
| ornitina | <0,01 | |
| lisina | <0,01 | |
| ácido aspártico | 7,7 | 13 |
| ácido glutâmico | 1,7 | 20 |
| serina | 1,1 | |
| treonina | 0,2 | |
| alotreonina | 0,2 | |
| metionina | 0,1 | |
| homocisteína | 0,5 | |
| homosserina | 0,5 | |
| β-alanina | 4,3 | 10 |
| ácido β-amino-n-butírico | 0,1 | 5 |
| ácido β-aminoisobutírico | 0,5 | 7 |
| ácido γ-aminobutírico | 0,5 | 7 |
| ácido α-aminoisobutírico | 7 | 33 |
| isovalina | 1 | 11 |
| sarcosina | 12,5 | 7 |
| N-etil glicina | 6,8 | 6 |
| N-propil glicina | 0,5 | |
| N-isopropil glicina | 0,5 | |
| N-metil alanina | 3,4 | 3 |
| N-etil alanina | <0,05 | |
| N-metil β-alanina | 1.0 | |
| N-etil β-alanina | <0,05 | |
| isoserina | 1,2 | |
| ácido α-hidroxi-γ-aminobutírico | 17 |
Corrente lateral reta
O código genético foi descrito como um acidente congelado e as razões pelas quais há apenas um aminoácido padrão de cadeia linear (alanina) podem ser simplesmente redundância com valina, leucina e isoleucina. No entanto, é relatado que os aminoácidos de cadeia direta formam hélices alfa muito mais estáveis.

Glicina (cadeia lateral de hidrogênio)
Alanina (cadeia lateral de metil)

Homoalanina ou ácido α-aminobutírico (cadeia lateral de etila)

Norvalina ( cadeia lateral de n- Propil)

Norleucina ( cadeia lateral de n- butil)

Homonorleucina ( cadeia lateral de n- pentil) (ácido heptanóico mostrado)




Calcogênio
Serina, homoserina , O-metil-homoserina e O-etil-homoserina possuem uma cadeia lateral de hidroximetil, hidroxietil, O-metil-hidroximetil e O-metil-hidroxietil. Enquanto a cisteína, homocisteína , metionina e etionina possuem os equivalentes de tiol. Os equivalentes do selenol são selenocisteína, selenohomocisteína, selenometionina e selenoetionina. Os aminoácidos com o próximo calcogênio inativo também são encontrados na natureza: várias espécies, como Aspergillus fumigatus, Aspergillus terreus e Penicillium chrysogenum na ausência de enxofre, são capazes de produzir e incorporar nas proteínas telurocisteína e telurometionina .
A hidroxiglicina, um aminoácido com uma cadeia lateral de hidroxila, é altamente instável.
Código genético expandido
Funções
Em células, especialmente autótrofas, vários aminoácidos não proteinogênicos são encontrados como intermediários metabólicos. No entanto, apesar da flexibilidade catalítica das enzimas de ligação de PLP, muitos aminoácidos são sintetizados como ceto-ácidos ( por exemplo, 4-metil-2-oxopentanoato em leucina) e aminados na última etapa, mantendo assim o número de aminoácidos não proteinogênicos intermediários bastante baixos.
Ornitina e citrulina ocorrem no ciclo da ureia , parte do catabolismo de aminoácidos (veja abaixo).
Além do metabolismo primário, vários aminoácidos não proteinogênicos são precursores ou a produção final no metabolismo secundário para fazer pequenos compostos ou peptídeos não ribossomais (como algumas toxinas ).
Incorporado pós-tradução na proteína
Apesar de não serem codificados pelo código genético como aminoácidos proteinogênicos, alguns aminoácidos não padronizados são encontrados nas proteínas. Estes são formados por modificação pós-tradução das cadeias laterais de aminoácidos padrão presentes na proteína alvo. Essas modificações são freqüentemente essenciais para a função ou regulação de uma proteína; por exemplo, no gama-carboxiglutamato a carboxilação do glutamato permite uma melhor ligação dos cátions de cálcio , e na hidroxiprolina a hidroxilação da prolina é crítica para a manutenção dos tecidos conjuntivos . Outro exemplo é a formação de hipusina no fator de iniciação da tradução EIF5A , por meio da modificação de um resíduo de lisina. Essas modificações também podem determinar a localização da proteína, por exemplo, a adição de grupos hidrofóbicos longos pode fazer com que uma proteína se ligue a uma membrana fosfolipídica .

Ácido carboxiglutâmico. Enquanto o ácido glutâmico possui um grupo γ-carboxila, o ácido carboxiglutâmico possui dois.

Hidroxiprolina. Este iminoácido difere da prolina devido a um grupo hidroxila no carbono 4.
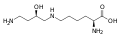
Hypusine . Este aminoácido é obtido adicionando ao grupo ε-amino de uma lisina uma fração 4-aminobutil (obtida a partir da espermidina )


-Pyroglutamic_acid_Structural_Formulae.png)
Existem algumas evidências preliminares de que o ácido aminomalônico pode estar presente, possivelmente por incorporação incorreta, na proteína.
Análogos tóxicos
Vários aminoácidos não proteinogênicos são tóxicos devido à sua capacidade de mimetizar certas propriedades de aminoácidos proteinogênicos, como a tialisina . Alguns aminoácidos não proteinogênicos são neurotóxicos por imitar aminoácidos usados como neurotransmissores (ou seja, não para a biossíntese de proteínas), por exemplo , ácido quisquálico , canavanina ou ácido azetidina-2-carboxílico . A cefalosporina C tem uma estrutura de ácido α-aminoadípico (homoglutamato) que é amidada com uma porção de cefalosporina. A penicilamina é um aminoácido terapêutico, cujo modo de ação é desconhecido.

Tialisina

Ácido quisquálico

Canavanina

ácido azetidina-2-carboxílico

Cefalosporina C

Penicilamina

-Azetidine-2-carboxylate.png)

As cianotoxinas de ocorrência natural também podem incluir aminoácidos não proteinogênicos. Microcistina e nodularina , por exemplo, são ambas derivadas de ADDA , um β-aminoácido.
Não são aminoácidos
A taurina é um ácido aminossulfônico e não um ácido aminocarboxílico, no entanto, ocasionalmente é considerada como tal, uma vez que as quantidades necessárias para suprimir o auxotrófico em certos organismos (por exemplo, gatos) são mais próximas dos "aminoácidos essenciais" (auxotrofia de aminoácidos) do que de vitaminas (auxotrofia de cofator).
Os osmólitos, sarcosina e glicina betaína são derivados de aminoácidos, mas possuem uma amina secundária e quaternária, respectivamente.