
Anatomia do cerebelo - Anatomy of the cerebellum
| Cerebelo | |
|---|---|
 Desenho do cérebro humano, mostrando cerebelo e ponte
| |

Seção transversal vertical da linha média do cerebelo humano, mostrando o padrão de dobramento do córtex e estruturas internas
| |
| Detalhes | |
| Parte de | Metencéfalo |
| Artéria | SCA , AICA , PICA |
| Veia | superior , inferior |
| Identificadores | |
| NeuroLex ID | birnlex_1489 |
| TA98 | A14.1.07.001 |
| TA2 | 5788 |
| Termos anatômicos de neuroanatomia | |
A anatomia do cerebelo pode ser visualizada em três níveis. No nível da anatomia macroscópica , o cerebelo consiste em uma camada de córtex fortemente dobrada e enrugada, com substância branca por baixo, vários núcleos profundos embutidos na substância branca e um ventrículo cheio de líquido no meio. No nível intermediário, o cerebelo e suas estruturas auxiliares podem ser divididos em várias centenas ou milhares de módulos ou compartimentos que funcionam independentemente, conhecidos como microzonas . No nível microscópico, cada módulo consiste no mesmo pequeno conjunto de elementos neuronais, dispostos com uma geometria altamente estereotipada.
Anatomia macroscópica
O cerebelo está localizado na base do cérebro, com a grande massa do córtex cerebral acima dele e a parte do tronco cerebral chamada ponte na frente dele. É separado do cérebro que o recobre por uma camada de dura-máter ; todas as suas conexões com outras partes do cérebro viajam através da ponte. Os anatomistas classificam o cerebelo como parte do metencéfalo , que também inclui a ponte; o metencéfalo, por sua vez, é a parte superior do rombencéfalo ou "rombencéfalo". Como o córtex cerebral, o cerebelo é dividido em dois hemisférios; ele também contém uma zona estreita da linha média chamada vermis . Um conjunto de grandes dobras é convencionalmente usado para dividir a estrutura geral em dez lóbulos menores .
Por causa de seu grande número de células granulares minúsculas , o cerebelo contém mais neurônios do que o resto do cérebro junto, mas ocupa apenas 10% do volume total do cérebro. O cerebelo recebe quase 200 milhões de fibras de entrada; em contraste, o nervo óptico é composto de apenas um milhão de fibras.
A aparência incomum da superfície do cerebelo esconde o fato de que a maior parte da estrutura é formada por uma camada de matéria cinzenta fortemente dobrada, o córtex cerebelar . Foi estimado que se o córtex cerebelar humano pudesse ser completamente desdobrado, daria origem a uma camada de tecido neural de cerca de 1 metro de comprimento e 10 centímetros de largura - uma área de superfície total de 500-1000 cm quadrados, todos embalados em um volume de 100-150 cm cúbicos. Debaixo da matéria cinzenta do córtex encontra-se a matéria branca , composta em grande parte por fibras nervosas mielinizadas indo e vindo do córtex. Embutidos na matéria branca - que às vezes é chamada de arbor vitae (Árvore da Vida) no cerebelo por causa de sua aparência ramificada em forma de árvore - estão quatro núcleos cerebelares profundos .
O cerebelo pode ser dividido de acordo com três critérios diferentes: macroscópico, filogenético e funcional.
Divisões anatômicas brutas
Na inspeção geral, três lobos podem ser distinguidos no cerebelo: o lobo floculonodular , o lobo anterior (rostral à "fissura primária") e o lobo posterior (dorsal à "fissura primária"). Os dois últimos podem ser divididos em um vermis cerebelar da linha média e hemisférios cerebelares laterais.

Figura 3: Cerebelo e regiões circundantes; vista sagital de um hemisfério. R: Midbrain . B: Pons . C: Medula . D: Medula espinhal . E: Quarto ventrículo . F: Arbor vitae . G: Amígdala . H: Lobo anterior. I: lobo posterior.
|

Divisões filogenéticas e funcionais
O cerebelo também pode ser dividido em três partes com base em critérios filogenéticos (a idade evolutiva de cada parte) e em critérios funcionais (as conexões de entrada e saída de cada parte e o papel desempenhado na função cerebelar normal). Do filogeneticamente mais antigo ao mais novo, as três partes são:
| Denominação funcional ( denominação filogenética) | Partes anatômicas | Função |
| Vestibulocerebelo (Arquicerebelo) | Lobo floculonodular (e vermis imediatamente adjacente) | O vestibulocerebelo regula o equilíbrio e os movimentos dos olhos. Ele recebe entrada vestibular de ambos os canais semicirculares e dos núcleos vestibulares e envia as fibras de volta para os núcleos vestibulares medial e lateral. Ele também recebe informações visuais dos colículos superiores e do córtex visual (este último via núcleos pontinos , formando uma via córtico-ponto-cerebelar). Lesões do vestibulocerebelo causam distúrbios de equilíbrio e marcha . Existe outra pequena região, conhecida como lóbulo biventro . |
| Espinocerebelo (Paleocerebelo) | Vermis e partes intermediárias dos hemisférios ("paravermis") | O espinocerebelo regula os movimentos do corpo e dos membros. Ele recebe dados de propriocepção das colunas dorsais da medula espinhal (incluindo o trato espinocerebelar ) e do nervo trigêmeo , bem como dos sistemas visual e auditivo . Ele envia fibras para núcleos cerebelares profundos (incluindo o núcleo fastigial ) que por sua vez se projetam para o córtex cerebral (via mesencéfalo e tálamo ) e para o tronco encefálico (via formação reticular na ponte e núcleos vestibulares na medula oblonga ), assim fornecendo modulação de sistemas motores descendentes. O espinocerebelo contém mapas sensoriais à medida que recebe dados sobre a posição de várias partes do corpo no espaço: em particular, o vermis recebe fibras do tronco e porções proximais dos membros, enquanto as partes intermediárias dos hemisférios recebem fibras das porções distais dos membros . O espinocerebelo é capaz de elaborar input proprioceptivo para antecipar a posição futura de uma parte do corpo durante o movimento, de forma "feed forward". |
| Cerebrocerebellum (Neocerebellum, Pontocerebellum) | Partes laterais dos hemisférios | O neocerebelo está envolvido no planejamento do movimento e na avaliação das informações sensoriais para a ação. Ele recebe entrada exclusivamente do córtex cerebral (especialmente o lobo parietal ) através dos núcleos pontinos (na ponte , formando vias cortico-ponto-cerebelares) e núcleo dentado (no cerebelo ), e envia fibras principalmente para o tálamo ventrolateral (em por sua vez conectado a áreas motoras do córtex pré-motor e área motora primária do córtex cerebral) e ao núcleo vermelho (por sua vez conectado ao núcleo olivar inferior , que se liga de volta aos hemisférios cerebelares). O neocerebelo está envolvido no planejamento do movimento que está prestes a ocorrer e também tem funções puramente cognitivas. |
Muito do que se compreende sobre as funções do cerebelo deriva da documentação cuidadosa dos efeitos das lesões focais em pacientes humanos que sofreram lesões ou doenças ou através da pesquisa de lesões em animais.
Anatomia celular
Conforme explicado com mais detalhes na seção Função , o cerebelo difere da maioria das outras áreas do cérebro porque o fluxo de sinais neurais por ele é quase inteiramente unidirecional: não há virtualmente nenhuma conexão retroativa entre seus elementos neuronais. Portanto, a maneira mais lógica de descrever a estrutura celular é começar com as entradas e seguir a sequência de conexões até as saídas.
Núcleos profundos
Os quatro núcleos profundos do cerebelo são os núcleos dentado , emboliforme , globoso e fastigii e atuam como os principais centros de comunicação, enviando e recebendo informações de e para partes específicas do cérebro. Além disso, esses núcleos recebem sinais inibitórios e excitatórios de outras partes do cérebro que, por sua vez, afetam os sinais de saída dos núcleos (os núcleos globoso e emboliforme constituem o núcleo interposto ).
Camadas corticais


A citoarquitetura ( organização celular ) do cerebelo é altamente uniforme, com conexões organizadas em uma matriz tridimensional de elementos de circuito perpendiculares . Essa uniformidade organizacional torna o circuito nervoso relativamente fácil de estudar.
Existem três camadas no córtex cerebelar; da camada externa para a interna, são as camadas molecular, de Purkinje e granular. A função do córtex cerebelar é essencialmente modular a informação que flui através dos núcleos profundos. O microcircuito do cerebelo é esquematizado na Figura 5. Fibras musgosas e trepadeiras transportam informações sensório-motoras para os núcleos profundos, que por sua vez as transmitem para várias áreas pré-motoras, regulando assim o ganho e o tempo das ações motoras. Fibras cobertas de musgo e trepadeiras também alimentam essas informações no córtex cerebelar, que realiza vários cálculos, resultando na regulação do disparo das células de Purkinje. Os neurônios de Purkinje realimentam os núcleos profundos por meio de uma sinapse inibitória potente . Essa sinapse regula até que ponto as fibras musgosas e trepadeiras ativam os núcleos profundos e, assim, controlam o efeito final do cerebelo na função motora. Foi demonstrado que a força sináptica de quase todas as sinapses no córtex cerebelar sofre plasticidade sináptica . Isso permite que os circuitos do córtex cerebelar ajustem e sintonizem continuamente a saída do cerebelo, formando a base de alguns tipos de aprendizagem e coordenação motora. Cada camada do córtex cerebelar contém os vários tipos de células que compõem esse circuito.
Camada molecular
Essa camada mais externa do córtex cerebelar contém dois tipos de interneurônios inibitórios : as células estreladas e as células em cesta . Ele também contém as árvores dendríticas dos neurônios de Purkinje e tratos de fibras paralelas das células granulares. As células estreladas e em cesta formam sinapses GABAérgicas com os dendritos das células de Purkinje.
Camada Purkinje
A camada do meio contém apenas um tipo de corpo celular - o da grande célula de Purkinje . As células de Purkinje são os neurônios integrativos primários do córtex cerebelar e fornecem sua única saída. Os dendritos das células de Purkinje são grandes árvores com centenas de ramos espinhosos alcançando a camada molecular (Fig. 6). Essas árvores dendríticas são planas - quase todas em planos - com as árvores Purkinje vizinhas em planos paralelos. Cada fibra paralela das células granulares passa ortogonalmente por essas árvores, como um fio que passa por muitas camadas. Os neurônios de Purkinje são gabaérgicos - o que significa que têm sinapses inibitórias - com os neurônios dos núcleos cerebelares profundos e vestibulares do tronco cerebral. Cada célula de Purkinje recebe estímulos excitatórios de 100.000 a 200.000 fibras paralelas. Diz-se que as fibras paralelas são responsáveis pelo spiking simples (tudo ou nada, invariante de amplitude ) da célula de Purkinje.
As células de Purkinje também recebem informações do núcleo olivar inferior por meio de fibras de escalada . Um bom mnemônico para essa interação é a frase "subir na outra oliveira", visto que as fibras trepadeiras se originam da oliveira inferior contralateral. Em notável contraste com as mais de 100.000 entradas de fibras paralelas, cada célula de Purkinje recebe entrada de exatamente uma fibra de escalada; mas essa fibra única "sobe" os dendritos da célula de Purkinje, enrolando-se em torno deles e fazendo um grande número de sinapses à medida que avança. A entrada líquida é tão forte que um único potencial de ação de uma fibra de escalada é capaz de produzir um "pico complexo" na célula de Purkinje: uma explosão de vários picos consecutivos, com amplitude decrescente, seguida por uma pausa durante a qual picos simples são suprimidos.
Logo abaixo da camada de Purkinje estão as células Lugaro, cujos dendritos muito longos viajam ao longo da fronteira entre o Purkinje e as camadas granulares.
Camada granular
A camada mais interna contém os corpos celulares de três tipos de células: as numerosas e minúsculas células granulares , as células em escova unipolares ligeiramente maiores e as células de Golgi muito maiores . Fibras musgosas entram na camada granular de seu principal ponto de origem, os núcleos pontinos. Essas fibras formam sinapses excitatórias com as células granulares e com as células dos núcleos cerebelares profundos. As células granulares enviam seus axônios em forma de T - conhecidos como fibras paralelas - para a camada molecular superficial, onde formam centenas de milhares de sinapses com dendritos de células de Purkinje . O cerebelo humano contém cerca de 60 a 80 bilhões de células granulares, tornando esse tipo de célula única , de longe, o neurônio mais numeroso do cérebro (cerca de 70% de todos os neurônios do cérebro e da medula espinhal, combinados). As células de Golgi fornecem feedback inibitório às células granulares, formando uma sinapse com elas e projetando um axônio na camada molecular.
Relação com o córtex cerebral
Os potenciais de campo locais do neocórtex e cerebelo oscilam coerentemente em (6–40 Hz) em animais que se comportam acordados. Eles parecem estar sob o controle da produção do córtex cerebral. Essa saída seria mediada por um caminho dos neurônios da camada 5/6 no neocórtex através desse projeto até a ponte ou a oliva inferior. Se através da ponte, isso iria para as fibras musgosas que fazem sinapses com os grânulos e os neurônios de Golgi com as células dos grânulos, então direcionando os neurônios de Purkinje por meio de suas fibras excitatórias paralelas. Se a azeitona inferior fosse transmitida por fibras de escalada excitatória para os neurônios de Purkinje. Estes retornam esta saída de volta ao córtex cerebral através do tálamo ventrolateral, completando a alça.
A via corticopontocerebelar é a maior via associada ao cerebelo. Surgindo no córtex cerebral, essas fibras terminam primeiro ipsilateralmente nos núcleos pontinos . Em seguida, as fibras decussam e formam o pedúnculo cerebelar médio, terminando no córtex cerebelar como fibras musgosas. Essa via transmite sinais que informam o cerebelo sobre o movimento em andamento e o movimento que se aproxima. Isso ajuda no ajuste contínuo da atividade motora.
O início do movimento é retransmitido para o cerebelo pela via corticorreticulocerebelar. Essas sinapses ipsilateralmente na formação reticular e , em seguida, através dos pedúnculos inferior e médio no vérmis cerebelar .
O córtex motor e o córtex somatossensorial se projetam nos núcleos olivares ipsilaterais inferior e acessório, formando então o trato olivocerebelar . As fibras cortico-olivares fazem sinapse bilateralmente no núcleo olivar inferior . A ordem é preservada nas projeções do trato olivocerebelar nos 'mapas corporais' no córtex cerebelar contralateral. Em condições de repouso em experimentos com animais, grupos de neurônios olivares descarregam sincronicamente em 5 a 10 Hz (impulsos / s). No córtex cerebelar, a resposta das células de Purkinje assume a forma de pontas complexas.
O cerebelo envia suas projeções de volta ao córtex cerebral através do trato cerebelotalâmico .
A expansão lateral cerebelar, ou neocerebelo, pode estar associada a funções cognitivas e está anatomicamente ligada ao córtex pré-frontal lateral . Apresenta maior atividade durante a fala, com predomínio unilateral consistente com uma possível ligação (via tálamo) com a área motora da fala.
Quando ocorrem lesões nas áreas de associação ligadas ao cerebelo por fibras corticopontocerebelares, pode ocorrer a síndrome cognitivo-afetiva. Isso resulta em defeitos cognitivos na forma de diminuição do poder de raciocínio, desatenção, erros gramaticais na fala, mau senso espacial e perda irregular de memória.
Fornecimento de sangue


Três artérias fornecem sangue ao cerebelo (Fig. 7): a artéria cerebelar superior (SCA), a artéria cerebelar inferior anterior (AICA) e a artéria cerebelar inferior posterior (PICA).
A SCA ramifica-se na porção lateral da artéria basilar, imediatamente inferior à sua bifurcação na artéria cerebral posterior . Aqui, ele se enrola posteriormente em torno da ponte (para a qual também fornece sangue) antes de chegar ao cerebelo. A SCA fornece sangue à maior parte do córtex cerebelar, aos núcleos cerebelares e aos pedúnculos cerebelares superiores.
O AICA ramifica-se na porção lateral da artéria basilar, imediatamente superior à junção das artérias vertebrais. Desde sua origem, ela se ramifica ao longo da porção inferior da ponte no ângulo cerebelopontino antes de atingir o cerebelo. Essa artéria fornece sangue à porção anterior do cerebelo inferior, ao pedúnculo cerebelar médio e aos nervos facial (NC VII) e vestibulococlear (NC VIII). A obstrução da AICA pode causar paresia , paralisia e perda de sensibilidade na face; também pode causar deficiência auditiva . Além disso, pode causar infarto do ângulo cerebelopontino. Isso poderia levar à hiperacusia (disfunção do músculo estapédio , inervado pelo NC VII ) e vertigem (interpretação errada da aceleração da endolinfa do canal semicircular vestibular causada pela alteração do NC VIII ).
O PICA ramifica-se na porção lateral das artérias vertebrais imediatamente inferiores à sua junção com a artéria basilar. Antes de atingir a superfície inferior do cerebelo, o PICA envia ramos para a medula, fornecendo sangue a vários núcleos de nervos cranianos . No cerebelo, o PICA fornece sangue para a porção posterior inferior do cerebelo, o pedúnculo cerebelar inferior, o núcleo ambíguo , o núcleo motor do vago , o núcleo do trigêmeo espinhal , o núcleo solitário e os núcleos vestibulococleares .
Variações entre vertebrados
Existe uma variação considerável no tamanho e na forma do cerebelo em diferentes espécies de vertebrados. Geralmente é maior em peixes cartilaginosos e ósseos , pássaros e mamíferos, mas um pouco menor em répteis. Os grandes lobos emparelhados e convolutos encontrados em humanos são típicos de mamíferos, mas o cerebelo é geralmente um único lobo mediano em outros grupos e é liso ou apenas ligeiramente sulcado. Em mamíferos, o neocerebelo é a maior parte do cerebelo em massa, mas em outros vertebrados, é normalmente o espinocerebelo.
Em anfíbios , lampreias e peixes - bruxa, o cerebelo é pouco desenvolvido; nos últimos dois grupos, mal se distingue do tronco cerebral. Embora o espinocerebelo esteja presente nesses grupos, as estruturas primárias são pequenos núcleos pareados que correspondem ao vestibulocerebelo.
Pedúnculos
O cerebelo segue o padrão geral de grupos de três encontrados na anatomia, com três pedúnculos cerebelares de entrada e saída principais (feixes de fibras). Estes são os pedúnculos cerebelares superior (brachium conjuntivum), médio (brachium pontis) e inferior ( corpos restiformes e justarestiformes ).
| Pedúnculo | Descrição |
| Superior | Embora existam algumas fibras aferentes do trato espinocerebelar anterior que são transportadas para o lobo cerebelar anterior por meio desse pedúnculo, a maioria das fibras são eferentes. Assim, o pedúnculo cerebelar superior é a principal via de saída do cerebelo. A maioria das fibras eferentes se origina no núcleo denteado que, por sua vez, se projeta para várias estruturas do mesencéfalo , incluindo o núcleo vermelho , o núcleo ventral lateral / ventral anterior do tálamo e a medula . As vias dentatorubrotalamocortical (núcleo dentado> núcleo vermelho > tálamo > córtex pré-motor ) e cerebelotalamocortical (cerebelo> tálamo> córtex pré-motor) são duas vias principais que passam por este pedúnculo e são importantes no planejamento motor . |
| Meio | Este é composto inteiramente de fibras aferentes originadas dentro dos núcleos pontinos como parte do trato corticopontocerebelar maciço (córtex cerebral> ponte> cerebelo). Essas fibras descendem das áreas sensoriais e motoras do neocórtex cerebral e tornam o pedúnculo cerebelar médio o maior dos três pedúnculos cerebelares. |
| Inferior | Isso carrega muitos tipos de fibras de entrada e saída que se preocupam principalmente com a integração da entrada sensorial proprioceptiva com as funções vestibulares motoras , como equilíbrio e manutenção da postura. As informações proprioceptivas do corpo são transportadas para o cerebelo por meio do trato espinocerebelar dorsal . Este trato passa pelo pedúnculo cerebelar inferior e faz sinapses dentro do paleocerebelo. Informações vestibulares são projetadas no arquicerebelo. As fibras trepadeiras da azeitona inferior percorrem o pedúnculo cerebelar inferior. Este pedúnculo também carrega informações diretamente das células de Purkinje para os núcleos vestibulares no tronco cerebral dorsal, localizado na junção entre a ponte e a medula . |
Existem três fontes de entrada para o cerebelo, em duas categorias consistindo de fibras musgosas e trepadeiras, respectivamente. Fibras musgosas podem se originar dos núcleos pontinos, que são agrupamentos de neurônios localizados na ponte que carregam informações do córtex cerebral contralateral. Eles também podem surgir dentro do trato espinocerebelar cuja origem está localizada na medula espinhal ipsilateral . A maior parte da saída do cerebelo inicialmente faz sinapses com os núcleos cerebelares profundos antes de sair pelos três pedúnculos. A exceção mais notável é a inibição direta dos núcleos vestibulares pelas células de Purkinje.
Desenvolvimento
Durante os primeiros estágios do desenvolvimento embrionário , o cérebro começa a se formar em três segmentos distintos: o prosencéfalo , o mesencéfalo e o rombencéfalo . O rombencéfalo é o segmento mais caudal (em direção à cauda) do cérebro embrionário; é a partir desse segmento que o cerebelo se desenvolve. Ao longo do segmento rombencefálico embrionário desenvolvem oito inchaços, chamados rombômeros . O cerebelo se origina de dois rombômeros localizados na placa alar do tubo neural , uma estrutura que acaba formando o cérebro e a medula espinhal. Os rombômeros específicos a partir dos quais o cerebelo se forma são o rombômero 1 (Rh.1) caudalmente (próximo à cauda) e o "istmo" rostralmente (próximo à frente).
Acredita-se que duas regiões primárias dêem origem aos neurônios que constituem o cerebelo. A primeira região é a zona ventricular no teto do quarto ventrículo . Essa área produz células de Purkinje e neurônios nucleares cerebelares profundos . Essas células são os neurônios de saída primários do córtex cerebelar e do cerebelo. A segunda zona germinativa (local de nascimento celular) é conhecida como lábio rômbico, os neurônios então se movem na semana embrionária humana 27 para a camada granular externa . Essa camada de células - encontrada no exterior do cerebelo - produz os neurônios granulares. Os neurônios granulares migram desta camada externa para formar uma camada interna conhecida como camada granular interna. A camada granular externa deixa de existir no cerebelo maduro, deixando apenas células granulares na camada granular interna. A substância branca cerebelar pode ser uma terceira zona germinativa no cerebelo; no entanto, sua função como zona germinativa é controversa.
Imagens adicionais
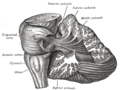
Dissecção mostrando as fibras de projeção do cerebelo
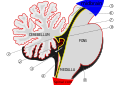
Esquema de teto do quarto ventrículo. A flecha está no forame de Majendie.

Visão mediana do cérebro humano
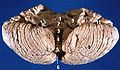
Vista anterior do cerebelo humano, com números indicando pontos de referência salientes



